Vier Astronauten werden mit Artemis II, dem ersten bemannten Flug im Rahmen der NASA-Artemis-Mission, den Mond umrunden und zurückkehren. Die NASA-Astronauten Reid Wiseman, Victor Glover und Christina Koch sowie der Astronaut der kanadischen Raumfahrtbehörde CSA, Jeremy Hansen, werden die ersten Menschen an Bord der SLS-Rakete (Space Launch System), des Orion-Raumschiffs und der dazugehörigen Bodensysteme sein, wenn die Crew in die lebensfeindliche Umgebung des Weltraums vordringt. Dieser Flug ist ein weiterer Schritt hin zu bemannten Missionen zur Mondoberfläche und hilft der NASA, sich auf zukünftige Astronautenmissionen zum Mars vorzubereiten.
Die eingereichten Namen werden auf einer SD-Karte gespeichert, die beim Start der Artemis-II-Mission im Jahr 2026 an Bord von Orion mitfliegen wird. Bleiben Sie auf dem Laufenden über die neuesten Missions-Updates, einschließlich Informationen zum Start, auf nasa.gov oder abonnieren Sie den Newsletter, um NASA-Updates direkt in Ihr Postfach zu erhalten.
Die NASA entsendet Astronauten zum Mond, um wissenschaftliche Erkenntnisse zu gewinnen, wirtschaftliche Vorteile zu erzielen und die Grundlage für die ersten bemannten Marsmissionen zu schaffen. Artemis II wird die Fähigkeiten der NASA im Weltraum testen, da erstmals Menschen mit der SLS-Rakete und dem Orion-Raumschiff fliegen. Die etwa zehntägige Mission startet spätestens im April 2026 vom Startkomplex 39 des Kennedy Space Center der NASA in Florida.
Die Besatzung wird in den ersten beiden Missionstagen erste Systemprüfungen an Orion durchführen und die Manövrierfähigkeit des Raumschiffs in Erdnähe manuell testen, bevor sie Kurs auf den Mond nimmt.
Das Servicemodul von Orion wird den nötigen Schub geben, um die Erdumlaufbahn zu verlassen und Kurs auf den Mond zu nehmen. Dieser translunare Impuls schickt die Astronauten auf eine viertägige Reise um die Mondrückseite in einer Achterbahn, die sich über 370.000 Kilometer von der Erde erstreckt. Am Ende der Reise fliegt die Besatzung etwa 7.400 Kilometer hinter dem Mond und testet dabei die Systeme des Raumschiffs. Mehrere Nutzlasten werden an Bord von Artemis II mitfliegen, um unser Wissen über Weltraumstrahlung, menschliche Gesundheit und Verhalten sowie Weltraumkommunikation zu erweitern. Die gewonnenen Erkenntnisse werden uns bei zukünftigen Erkundungsmissionen helfen.
Orion wird mit hoher Geschwindigkeit in die Erdatmosphäre eintreten und sicher im Pazifik vor der Küste von San Diego wassern. Dort wird ein Bergungsteam der NASA und des US-Verteidigungsministeriums die Besatzung und das Raumschiff bergen.
Eine Untersuchung von Wikingerskeletten enthüllt eine verborgene Geschichte von Gewalt, Macht und überraschenden Unterschieden zwischen benachbarten Wikingergesellschaften.
Die Knochen stammen aus ganz Norwegen – aus dem Norden und aus der Region des Oslofjords – und überraschend viele von ihnen wiesen Spuren brutaler Todesursachen auf.
Einige hatten Schädel, die durch stumpfe Gewalt zertrümmert wurden. Andere waren mit Schwertern in Rücken oder Beine geschlagen worden. In einigen steckten noch Pfeilspitzen.
„Die relativ wenigen menschlichen Überreste, die wir untersuchen konnten, zeugten von großer Gewalt“, sagte Lisa Strand, eine Osteologin, deren Untersuchung der Wikingerreste Teil ihrer Doktorarbeit an der NTNU war.
Insgesamt wiesen fast 40 Prozent der von ihr untersuchten Wikingerreste Hinweise auf einen gewaltsamen Tod auf.
Wie Strand und eine kleine Gruppe von Forschern herausfanden, stammten diese Wikingerknochen nicht nur von unglücklichen Individuen, sondern waren Teil eines Musters.
Und ein Schlüsselelement des Puzzles war, daß es sich um Wikinger aus dem heutigen Norwegen handelte. Fast alle dänischen Wikinger wurden hingerichtet, indem man ihnen den Kopf abschlug! So etwas tut man im Kampf nicht, sagte Strand.
„Es ist ziemlich zeitaufwendig, jemanden zu enthaupten, selbst im Kampf. Man nimmt sich nicht die Zeit, ihm den Kopf abzuschlagen. Nein, nein. Es ist ziemlich düster, aber es ist wahr“, sagte sie im Podcast.
Das war ein entscheidender Hinweis, denn es deutet darauf hin, daß es in Dänemark Menschen mit starken Anführern gab, die genug Macht hatten, eine Hinrichtung anzuordnen.
„Die Tatsache, daß wir in Dänemark hauptsächlich Hinrichtungen sehen, deutet auf eine stärker zentralisierte Autorität hin“, sagte David Jacobson, Soziologe der University of South Florida, der Teil des Forschungsteams war.
Das James-Webb-Weltraumteleskop der NASA hat neue Details der Polarlichter auf dem größten Planeten unseres Sonnensystems eingefangen. Die auf Jupiter beobachteten tanzenden Lichter sind hundertmal heller als die auf der Erde. Dank Webbs hoher Empfindlichkeit konnten Astronomen die Phänomene untersuchen, um Jupiters Magnetosphäre besser zu verstehen.
Polarlichter entstehen, wenn hochenergetische Teilchen in die Atmosphäre eines Planeten in der Nähe seiner Magnetpole eintreten und mit Atomen oder Gasmolekülen kollidieren. Auf der Erde sind diese Polarlichter als Nord- und Südlichter bekannt. Die Polarlichter auf Jupiter sind nicht nur riesig, sondern auch hundertmal energiereicher als die in der Erdatmosphäre. Die Polarlichter der Erde entstehen durch Sonnenstürme – wenn geladene Teilchen von der Sonne auf die obere Atmosphäre niedergehen, Gase mit Energie versorgen und sie in Rot-, Grün- und Violetttönen leuchten lassen.
Foto A: Nahaufnahme der Beobachtungen der Polarlichter des Jupiters
NASA, ESA, CSA, Jonathan Nichols (Universität Leicester), Mahdi Zamani (ESA/Webb)
Jupiter hat eine weitere Quelle für seine Polarlichter: Das starke Magnetfeld des Gasriesen fängt geladene Teilchen aus seiner Umgebung ein. Dazu gehören nicht nur die geladenen Teilchen des Sonnenwinds, sondern auch die Teilchen, die von seinem umkreisenden Mond Io, bekannt für seine zahlreichen und großen Vulkane, ins All geschleudert werden. Ios Vulkane spucken Teilchen aus, die der Schwerkraft des Mondes entkommen und Jupiter umkreisen. Auch eine Flut geladener Teilchen, die von der Sonne freigesetzt wird, erreicht den Planeten. Jupiters großes und starkes Magnetfeld fängt alle geladenen Teilchen ein und beschleunigt sie auf enorme Geschwindigkeiten. Diese schnellen Teilchen treffen mit hohen Energien auf die Atmosphäre des Planeten, wodurch das Gas angeregt und zum Leuchten gebracht wird.
Die einzigartigen Fähigkeiten des Webb-Teleskops liefern nun neue Einblicke in die Polarlichter des Jupiters. Die hohe Empfindlichkeit des Teleskops ermöglicht es Astronomen, schnell variierende Polarlichtmerkmale zu erfassen. Neue Daten wurden am 25. Dezember 2023 mit Webbs NIRCam (Nahinfrarotkamera) von einem Wissenschaftlerteam unter der Leitung von Jonathan Nichols von der Universität Leicester im Vereinigten Königreich erfasst.
„Was für ein Weihnachtsgeschenk! Ich war einfach hin und weg!“, erzählte Nichols. „Wir wollten sehen, wie schnell sich die Polarlichter verändern, und erwarteten, dass sie langsam, vielleicht über eine Viertelstunde, auf- und abschwellen würden. Stattdessen beobachteten wir, wie die gesamte Polarlichtregion vor Licht sprühte und knallte, manchmal sogar sekundenschnell.“
Das Team untersuchte insbesondere die Emission des Trihydrogenkations (H3+), das in Polarlichtern entstehen kann. Es stellte fest, daß diese Emission weitaus variabler ist als bisher angenommen. Die Beobachtungen werden dazu beitragen, das Verständnis der Erwärmung und Abkühlung der oberen Atmosphäre des Jupiters zu vertiefen.
Das Team entdeckte in seinen Daten außerdem einige ungeklärte Beobachtungen.
„Was diese Beobachtungen noch spezieller machte, war die Tatsache, daß wir gleichzeitig mit dem Hubble-Weltraumteleskop der NASA Bilder im Ultraviolettbereich machten“, fügte Nichols hinzu. „Seltsamerweise gab es für das hellste von Webb beobachtete Licht kein reales Gegenstück in Hubbles Bildern. Das hat uns vor Rätsel gestellt. Um die von Webb und Hubble beobachtete Helligkeitskombination zu erzeugen, bedarf es einer Kombination großer Mengen sehr energiearmer Teilchen, die auf die Atmosphäre treffen, was bisher für unmöglich gehalten wurde. Wir verstehen immer noch nicht, wie das passiert.“
Das Nancy Grace Roman Space Telescope, benannt nach der ersten Chefastronomin der NASA, der „Mutter des Hubble-Weltraumteleskops“, wird ein mindestens 100-mal größeres Sichtfeld als das Hubble-Teleskop haben und während seiner Lebensdauer möglicherweise Licht von einer Milliarde Galaxien messen können. Dieses Observatorium wird auch in der Lage sein, Sternenlicht zu blockieren, um Exoplaneten und planetenbildende Scheiben direkt zu sehen, eine statistische Zählung der Planetensysteme in unserer Galaxie durchzuführen und wesentliche Fragen in den Bereichen dunkle Energie, Exoplaneten und Infrarot-Astrophysik zu klären.
Forscher haben die bisher am weitesten entfernte, milchstraßenähnliche Galaxie entdeckt. Diese Scheibengalaxie mit dem Namen REBELS-25 scheint so geordnet wie heutige Galaxien, aber wir sehen sie so, wie sie war, als das Universum erst 700 Millionen Jahre alt war. Das ist überraschend, da nach unserem derzeitigen Verständnis der Galaxienbildung solche frühen Galaxien chaotischer erscheinen sollten. Die Rotation und Struktur von REBELS-25 wurden mithilfe des Atacama Large Millimeter/submillimeter Array (ALMA) entdeckt, an dem die Europäische Südsternwarte (ESO) beteiligt ist.
Die Galaxien, die wir heute sehen, haben sich weit von ihren chaotischen, klumpigen Gegenstücken entfernt, die Astronomen normalerweise im frühen Universum beobachten. „Nach unserem Verständnis der Galaxienbildung erwarten wir, daß die meisten frühen Galaxien klein und chaotisch aussehen“, sagt Jacqueline Hodge, Astronomin an der Universität Leiden in den Niederlanden und Co-Autorin der Studie.
Diese chaotischen, frühen Galaxien verschmelzen miteinander und entwickeln sich dann mit unglaublich langsamer Geschwindigkeit zu glatteren Formen. Aktuelle Theorien legen nahe, daß Milliarden von Jahren der Entwicklung vergangen sein müssen, damit eine Galaxie so geordnet ist wie unsere eigene Milchstraße – eine rotierende Scheibe mit ordentlichen Strukturen wie Spiralarmen. Die Entdeckung von REBELS-25 stellt diesen Zeitrahmen jedoch in Frage.
In der Studie, die zur Veröffentlichung in Monthly Notices of the Royal Astronomical Society angenommen wurde, fanden Astronomen heraus, daß REBELS-25 die am weitesten entfernte, stark rotierende Scheibengalaxie ist, die jemals entdeckt wurde. Das Licht, das uns von dieser Galaxie erreicht, wurde ausgestrahlt, als das Universum erst 700 Millionen Jahre alt war – gerade einmal fünf Prozent seines heutigen Alters (13,8 Milliarden Jahre) , was die geordnete Rotation von REBELS-25 unerwartet macht. „Eine Galaxie zu sehen, die so stark von Rotation dominiert wird und so große Ähnlichkeiten mit unserer Milchstraße aufweist, stellt unser Verständnis davon in Frage, wie schnell sich Galaxien im frühen Universum zu den geordneten Galaxien des heutigen Kosmos entwickelten“, sagt Lucie Rowland, Doktorandin an der Universität Leiden und Erstautorin der Studie.
REBELS-25 wurde erstmals in früheren Beobachtungen desselben Teams entdeckt, die ebenfalls mit ALMA durchgeführt wurden, das sich in der chilenischen Atacama-Wüste befindet. Damals war es eine aufregende Entdeckung, die Hinweise auf Rotation zeigte, aber die Auflösung der Daten war nicht fein genug, um sicher zu sein. Um die Struktur und Bewegung der Galaxie richtig zu erkennen, führte das Team Folgebeobachtungen mit ALMA mit einer höheren Auflösung durch, die ihren rekordverdächtigen Charakter bestätigten. „ALMA ist das einzige existierende Teleskop mit der Empfindlichkeit und Auflösung, um dies zu erreichen“, sagt Renske Smit, Forscherin an der Liverpool John Moores University in Großbritannien und ebenfalls Co-Autorin der Studie.
Überraschenderweise deuteten die Daten auch auf weiter entwickelte Strukturen hin, die denen der Milchstraße ähneln, wie einen zentralen, länglichen Balken und sogar Spiralarme, obwohl weitere Beobachtungen erforderlich sein werden, um dies zu bestätigen. „Weitere Beweise für weiter entwickelte Strukturen zu finden, wäre eine aufregende Entdeckung, da es sich um die am weitesten entfernte Galaxie mit derartigen Strukturen handeln würde, die bisher beobachtet wurde“, sagt Rowland.
Diese zukünftigen Beobachtungen von REBELS-25 werden zusammen mit anderen Entdeckungen früher rotierender Galaxien möglicherweise unser Verständnis der frühen Galaxienbildung und der Entwicklung des Universums als Ganzes verändern.
Weitere Informationen Diese Forschung wird in einem Artikel mit dem Titel „REBELS-25: Entdeckung einer dynamisch kalten Scheibengalaxie bei z=7,31“ vorgestellt, der in den Monthly Notices of the Royal Astronomical Society erscheinen wird.
Die Beobachtungen wurden im Rahmen des ALMA Large Program REBELS: Reionization Era Bright Emission Lines Survey durchgeführt.
Das Team besteht aus L. E. Rowland (Leiden Observatory, Universität Leiden, Niederlande [Leiden]), J. Hodge (Leiden), R. Bouwens (Leiden), P. M. Piña (Leiden), A. Hygate (Leiden), H. Algera (Astrophysical Science Center, Hiroshima University, Japan [HASC]; National Astronomical Observatory of Japan, Japan), M. Aravena (Núcleo de Astronomía, Facultad de. Ingen). iería y Ciencias, Universidad Diego Portales, Chile), R. Bowler (Jodrell Bank Centre for Astrophysics, University of Manchester, UK), E. da Cunha (International Centre for Radio Astronomy Research, University of Western Australia, Australien; ARC Centre of Excellence for All Sky Astrophysics in 3 Dimensions), P. Dayal (Kapteyn Astronomical Institute, University of Groningen, Niederlande), A. Ferrara (Scuola Normale Superiore, Italien [SNS]), T. Herard-Demanche (Leiden), H. Inami (HASC), I. van Leeuwen (Leiden), I. de Looze (Sterrenkundig Observatorium, Universität Gent, Belgien), P. Oesch (Abteilung für Astronomie, Universität Genf, Schweiz; Cosmic Dawn Center, Dänemark), A. Pallottini (SNS), S. Phillips (Astrophysics Research Institute, Liverpool John Moores University, UK [LJMU]), M. Rybak (Fakultät für Elektrotechnik, Technische Universität Delft, Niederlande; Leiden; Niederländisches Institut für Weltraumforschung, Niederlande), S. Schouws (Leiden), R. Smit (LJMU), L. Sommovigo (Center for Computational Astrophysics, Flatiron Institute, USA), M. Stefanon (Departament d’Astronomia i Astrofísica, Universitat de València, Spanien; Grupo de Astrofísica Extragaláctica y Cosmología, Universitat de València, Spanien), P. van der Werf (Leiden).
Das Atacama Large Millimeter/submillimeter Array (ALMA), eine internationale astronomische Einrichtung, ist eine Partnerschaft zwischen ESO, der amerikanischen National Science Foundation (NSF) und den National Institutes of Natural Sciences (NINS) Japans in Zusammenarbeit mit der Republik Chile. ALMA wird von ESO im Namen seiner Mitgliedsstaaten, von NSF in Zusammenarbeit mit dem National Research Council of Canada (NRC) und dem National Science and Technology Council (NSTC) in Taiwan und von NINS in Zusammenarbeit mit der Academia Sinica (AS) in Taiwan und dem Korea Astronomy and Space Science Institute (KASI) finanziert.
ALMA-Bau und -Betrieb werden von ESO im Namen seiner Mitgliedsstaaten geleitet; vom National Radio Astronomy Observatory (NRAO), das von Associated Universities, Inc. (AUI) verwaltet wird, im Namen Nordamerikas; und vom National Astronomical Observatory of Japan (NAOJ) im Namen Ostasiens. Das Joint ALMA Observatory (JAO) übernimmt die einheitliche Leitung und Verwaltung des Baus, der Inbetriebnahme und des Betriebs von ALMA. Die Europäische Südsternwarte (ESO) ermöglicht es Wissenschaftlern weltweit, die Geheimnisse des Universums zum Wohle aller zu lüften. Wir entwerfen, bauen und betreiben bodengestützte Observatorien von Weltklasse – mit denen Astronomen spannende Fragen angehen und die Faszination der Astronomie verbreiten – und fördern die internationale Zusammenarbeit im Bereich der Astronomie. Die 1962 als zwischenstaatliche Organisation gegründete ESO wird heute von 16 Mitgliedstaaten (Österreich, Belgien, Tschechien, Dänemark, Frankreich, Finnland, Deutschland, Irland, Italien, die Niederlande, Polen, Portugal, Spanien, Schweden, die Schweiz und das Vereinigte Königreich) sowie dem Gastgeberland Chile und Australien als strategischen Partner unterstützt. Der Hauptsitz der ESO sowie ihr Besucherzentrum und Planetarium, die ESO Supernova, befinden sich in der Nähe von München in Deutschland, während unsere Teleskope in der chilenischen Atacamawüste stehen, einem wunderbaren Ort mit einzigartigen Bedingungen zur Himmelsbeobachtung.
Die ESO betreibt drei Beobachtungsstandorte: La Silla, Paranal und Chajnantor. Auf dem Paranal betreibt die ESO das Very Large Telescope und sein Very Large Telescope Interferometer sowie Durchmusterungsteleskope wie VISTA. Ebenfalls auf dem Paranal wird die ESO das Cherenkov Telescope Array South beherbergen und betreiben, das größte und empfindlichste Gammastrahlenobservatorium der Welt. Gemeinsam mit internationalen Partnern betreibt die ESO ALMA auf Chajnantor, eine Einrichtung, die den Himmel im Millimeter- und Submillimeterbereich beobachtet. Auf dem Cerro Armazones in der Nähe des Paranal bauen wir „das größte Auge der Welt am Himmel“ – das Extremely Large Telescope der ESO. Von unseren Büros in Santiago, Chile, aus unterstützen wir unsere Aktivitäten im Land und arbeiten mit chilenischen Partnern und der Gesellschaft zusammen.
Nachdem ein NASA-Astronaut im Rahmen der SpaceX Crew-8-Mission der NASA am Freitag sicher auf der Erde gelandet war, trat bei ihm ein medizinisches Problem auf. Die NASA-Astronauten Matthew Dominick, Michael Barratt und Jeanette Epps sowie der Roscosmos-Kosmonaut Alexander Grebenkin wurden gemeinsam zum Ascension Sacred Heart Pensacola in Florida geflogen.
Nach der medizinischen Untersuchung im Krankenhaus verließen drei der Besatzungsmitglieder Pensacola und sind im Johnson Space Center der NASA in Houston angekommen.
Der eine Astronaut, der auf Ascension verbleibt, befindet sich in stabilem Zustand und wird vorsorglich beobachtet. Um die medizinische Privatsphäre des Besatzungsmitglieds zu schützen, werden keine spezifischen Details zum Zustand oder zur Identität des Einzelnen weitergegeben.
Bei seiner Rückkehr zur Erde führte die SpaceX Dragon einen normalen Eintritt und eine normale Wasserung durch. Die Bergung der Besatzung und des Raumschiffs verlief ohne Zwischenfälle. Während routinemäßiger medizinischer Untersuchungen auf dem Bergungsschiff wurde aus Vorsicht die zusätzliche Untersuchung der Besatzungsmitglieder angefordert.
Wir sind Ascension Sacred Heart für seine Unterstützung in dieser Zeit dankbar und stolz auf unser Team und sein schnelles Handeln, um die Sicherheit unserer Crewmitglieder zu gewährleisten.
Die Crew-8-Crewmitglieder wasserten an Bord ihres Dragon-Raumschiffs in der Nähe von Pensacola, Florida, um eine 235-tägige Mission abzuschließen, von der sie 232 Tage an Bord der Internationalen Raumstation verbrachten, um wissenschaftliche Forschung zu betreiben.
Die Teams von NASA und SpaceX haben den nächsten Starttermin für die SpaceX Crew-9-Mission der NASA auf frühestens 13:17 Uhr EDT am Samstag, den 28. September, von der Cape Canaveral Space Force Station in Florida verschoben, da in der Region tropische Stürme zu erwarten sind. Die Änderung ermöglicht es den Teams, am Dienstagabend eine Probe der Startaktivitäten mit dem SpaceX Dragon-Raumschiff und der Falcon 9-Rakete durchzuführen, die früher am Tag zum Space Launch Complex-40 gerollt waren. Nach den Probeaktivitäten wird das integrierte System vor möglichen Sturmaktivitäten in den Hangar zurückkehren.
Obwohl der tropische Sturm Helene durch den Golf von Mexiko zieht und voraussichtlich den Florida Panhandle treffen wird, ist das Sturmsystem groß genug, daß in den Regionen Cape Canaveral und Merritt Island an Floridas Ostküste mit starken Winden und heftigen Regenfällen zu rechnen ist.
NASA-Astronaut Nick Hague und Roscosmos-Kosmonaut Aleksandr Gorbunov werden an Bord des Raumschiffs Dragon zur Internationalen Raumstation starten. Dies wird die neunte Crew-Rotation-Mission mit SpaceX im Rahmen des Commercial Crew Program der NASA sein. Während ihrer fünfmonatigen Mission werden sie Forschungs- und Wartungsarbeiten durchführen. Der Start der Mission erfolgt vom Space Launch Complex-40 auf der Cape Canaveral Space Force Station in Florida.
Abstrakt Der größte Primat aller Zeiten und eine der größten der südostasiatischen Megafauna, Gigantopithecus blacki1, überlebte in China etwa 2,0 Millionen Jahre lang bis zum späten Mittelpleistozän, als er ausstarb2,3,4. Sein Untergang ist rätselhaft, wenn man bedenkt, daß er einer der wenigen asiatischen Menschenaffen war, die in den letzten 2,6 Millionen Jahren ausgestorben sind, während andere, darunter der Orang-Utan, bis heute überlebt haben5. Die Ursache für das Verschwinden von G. blacki bleibt ungeklärt, könnte aber Aufschluß über die Widerstandsfähigkeit der Primaten und das Schicksal der Megafauna in dieser Region geben6. Hier haben die Forscher drei multidisziplinäre Analysen – Zeitablauf, vergangene Umgebungen und Verhalten – auf 22 Höhlen in Südchina angewendet. Sie haben 157 radiometrische Alter aus sechs Datierungstechniken verwendet, um einen Zeitplan für das Ableben von G. blacki zu erstellen. Sie zeigen, daß die Umwelt vor 2,3 Millionen Jahren ein Mosaik aus Wäldern und Gräsern war, das ideale Bedingungen für gedeihende G. blacki-Populationen bot. Kurz vor und während des Aussterbefensters vor 295.000 bis 215.000 Jahren kam es jedoch aufgrund der zunehmenden Saisonalität zu einer erhöhten Umweltvariabilität, die zu Veränderungen in den Pflanzengemeinschaften und einer Zunahme offener Waldumgebungen führte. Obwohl es seinem nahen Verwandten Pongo weidenreichi gelang, seine Ernährungspräferenzen und sein Verhalten an diese Variabilität anzupassen, zeigte G. blacki Anzeichen von chronischem Stress und schrumpfenden Populationen. Letztendlich führte sein Anpassungskampf zum Aussterben des größten Primaten, der jemals auf der Erde gelebt hat.
Kerninhalt Unser derzeitiges Verständnis von Gigantopithecus blacki beruht auf Höhlenablagerungen des frühen bis mittleren Pleistozäns in Südchina zwischen dem Jangtsekiang und dem Südchinesischen Meer (Abb. 1 und Abschnitt 1 mit ergänzenden Informationen). Diese Pongine7 gilt als Schlüsselmitglied der früh- bis mittelpleistozänen Faunenzonen Gigantopithecus–Sinomastodon und Stegodon–Ailuropoda im (sub)tropischen Ostasien, von etwa 2,0 Millionen Jahren (Ma) bis 330 Tausend Jahren (ka) 2,3 ,8,9. Er ist bekannt für seine ungewöhnlich großen Backenzähne, seine atypische Schmelzdicke, seine geschätzte Körpergröße von etwa 3 m und sein Gewicht von 200–300 kg und ist damit der größte Primat, der jemals auf der Erde gelebt hat4. Trotz 85 jähriger Suche beschränkt sich der Fossilienbestand von G. blacki auf vier Mandibeln und fast 2.000 isolierte Zähne ohne postkranielle Beweise4. Seine erste Entdeckung in einer Apotheke in Hongkong als „Drachenzahn“1 löste eine Suche nach den ersten In-situ-Funden aus10 (Weiterführende Abbildung 1f) und gipfelte in der Entdeckung mehrerer Höhlenstandorte in zwei Hauptgebieten, Chongzuo und Bubing Becken, in der ZAR-Provinz Guangxi4. Diese Stätten enthalten entscheidende Beweise für sein Überleben und seinen späteren Untergang.
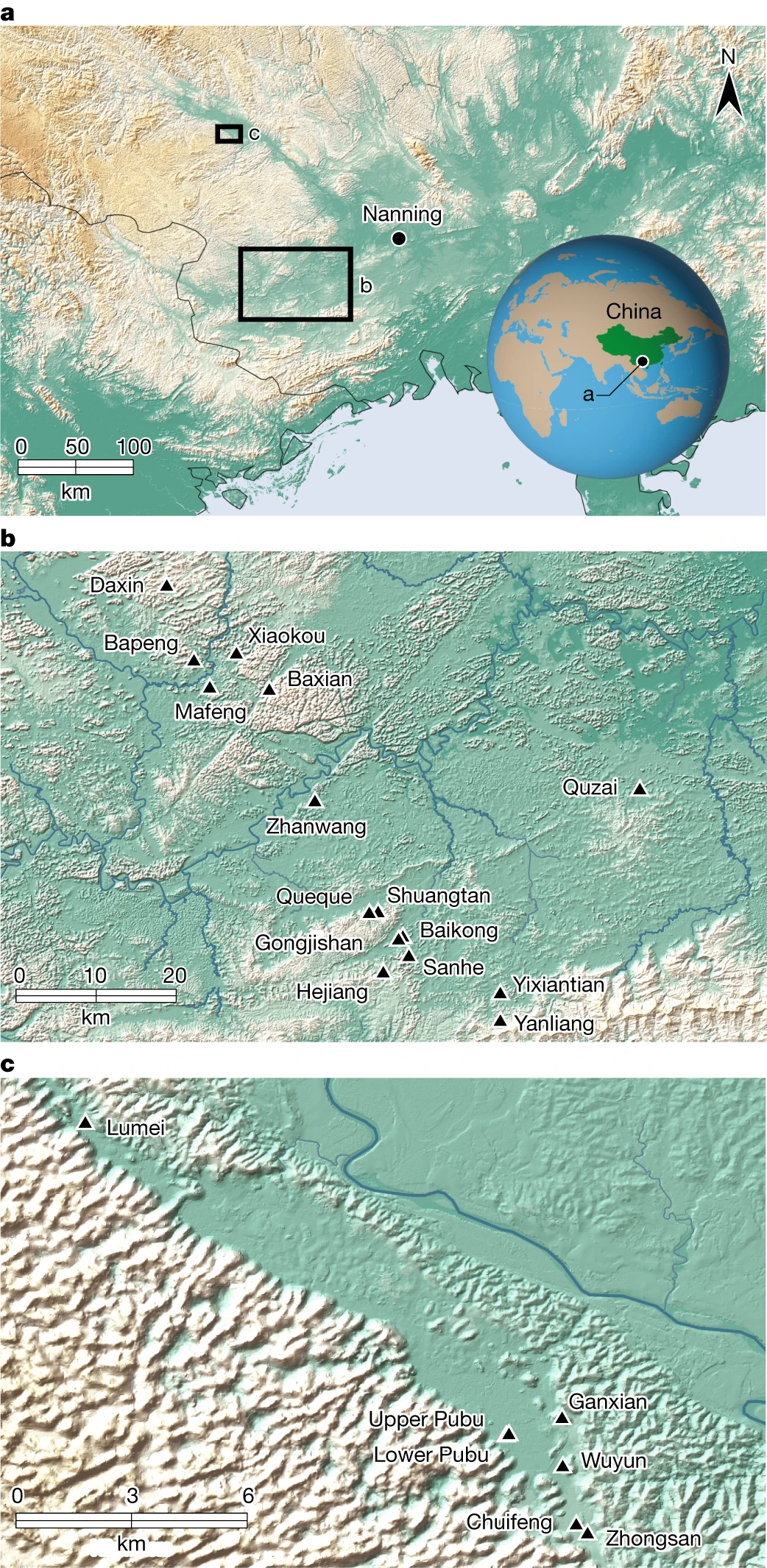
Fig. 1: Die Verortung der Studiengebiete dieser Forschungsergebnisse.
a–c, Die Lage von Südchina, der ZAR-Provinz Guangxi und der Stadt Nanning (a), wobei die Lage des Chongzuo-Untersuchungsgebiets durch ein großes Kästchen (b) und das Bubing-Becken-Untersuchungsgebiet durch ein kleineres Kästchen markiert ist ( C). b, Die Lage der 16 analysierten Höhlenstandorte im Chongzuo-Untersuchungsgebiet. c, Die Lage der sechs Höhlen, die im Untersuchungsgebiet des Bubing-Beckens analysiert wurden, einschließlich sowohl G. blacki-tragender als auch nicht-G. blacki-haltiger Höhlen. Blacki-haltige Höhlen aus beiden Regionen.
Nur sehr wenige dieser G. blacki-Standorte wurden mit mehr als einer radiometrischen Technik datiert; Daher bleibt der Zeitpunkt des Aussterbens ungewiss11. Die aktuelle Zeitspanne für sein Vorkommen liegt zwischen 2,2 Ma (Baikong-Höhle12) und 420–330 ka (Hejiang-Höhle9). Während dieser Zeit erfuhr G. blacki morphologische Veränderungen, einschließlich einer Zunahme der Zahngröße13 und der Zahnkomplexität9, was offenbar auf eine Ernährungsumstellung als Reaktion auf den ökologischen Druck hindeutet13. Rekonstruktionen der Ernährung von G. blacki auf der Grundlage der Zahnanatomie weisen auf einen spezialisierten Pflanzenfresser mit Anpassungen für den Verzehr abrasiver Nahrung14,15, starkes Kauen faseriger Nahrung16,17 und eine fruchtreiche Ernährung6,18 hin. Das vielfältige Waldökosystem zur Zeit von Baikong war in der Lage, die Biomasse mehrerer Primatengemeinschaften4 in einem weiten Gebiet in den Provinzen Guangxi, Guizhou, Hainan und Hubei zu unterstützen19. Zur Zeit von Hejiang war das Verbreitungsgebiet von G. blacki jedoch drastisch auf nur noch Guangxi9,13 zurückgegangen. Die Gründe für diesen dramatischen Rückgang und das letztendliche Aussterben sind nach wie vor heftig umstritten4, da es an einem regionalen Ansatz, einer Konzentration auf einzelne Standorte und Methoden und dem Fehlen von Verhaltens-4 und Umweltbeweisen20 mangelt.
Um die möglichen Ursachen für das Aussterben von G. blacki zu identifizieren, haben die Wissenschaftler einen regionalen Ansatz auf 22 Höhlen in Chongzuo und Bubing Basin angewendet, die entweder G. blacki-tragend (11) oder nicht-G. blacki-haltig waren. Blacki-haltige (11) Höhlenablagerungen (Erweiterte Daten, Abb. 1 und 2 und Ergänzende Informationen, Abschnitte 2 und 3). Mithilfe einer Kombination aus früheren Ausgrabungen (1999–2016) und neu entdeckten Höhlen (2017–2020) identifizierten und beprobten die Forscher fossile Brekzien für Datierungen, Paläoklima-Proxies und Verhaltensanalysen. Sie haben sechs unabhängige Datierungstechniken auf die Sedimente (Post-Infrarot-Infrarot-stimulierte Lumineszenz (pIR-IRSL), optisch stimulierte Lumineszenz (OSL), Elektronenspinresonanz (ESR) auf Quarz und U-Serie auf Speläothem) und Fossilien (U-Infrarot-stimulierte Lumineszenz) angewendet. Reihen zu Zähnen, gekoppelt US-ESR), um einen Bayes’schen modellierten Altersbereich für jeden Standort zu bestimmen (Ergänzende Informationen, Abschnitte 4–8), die dann weiter modelliert wurden, um ein regionales Aussterbefenster (EW) bereitzustellen. Sie haben Pollen-, Holzkohle-, paläontologische, stabile Isotopen- und mikrostratigraphische Analysen auf die Sedimente und Fossilien angewendet, um die früheren Umgebungen zu rekonstruieren (Ergänzende Informationen, Abschnitte 3 und 10–12). Schließlich haben wir eine Spurenelement-, stabile Isotopen- und Dental-Microwear-Texturanalyse (DMTA) auf die Zähne von G. blacki und den engsten Verwandten von Pongo weidenreichi angewendet, um etwaige Veränderungen in der Ernährung und im Verhalten von G. blacki vor und innerhalb der EW zu bestimmen, die damit in Zusammenhang stehen könnten bis zu seinem Untergang (Ergänzende Informationen, Abschnitte 12–14).
Den 157 radiometrischen Altersschätzungen zufolge liegen die Fossilfunde in den 22 Höhlen zwischen 2.300 und 49 ka (Abb. 2a und 3a, Erweiterte Daten Abb. 3–6 und Ergänzende Informationen in den Abschnitten 4–8 für alle Datierungstabellen und Diskussion von Einschränkungen). und Unsicherheiten). Diese Studie erweitert die Zeitachse für das Vorkommen von G. blacki von 2,3 Ma auf 255 ka, liefert einen genauen Zeitpunkt für das Fenster des Aussterbens bei 295–215 ka (2σ) (Ergänzende Informationen, Abschnitt 9) und legt Schwerpunkte für die Paläoumwelt und fest Verhaltensanalyse (Prä-EW (2.300–700 ka), Übergangsphase (700–295 ka), EW (295–215 ka) und Post-EW (215 ka bis heute)).
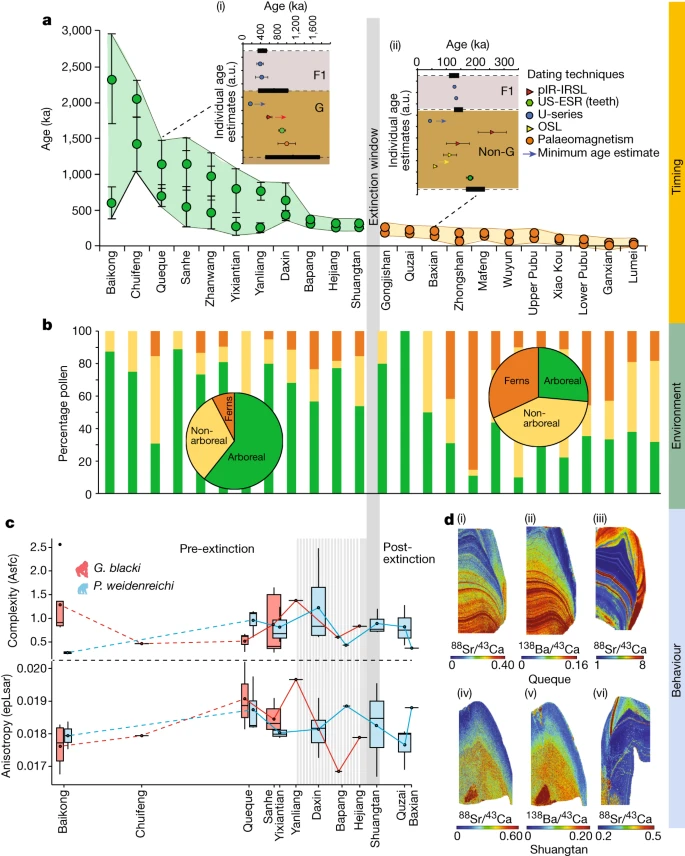
Fig. 2: Beispieldatensätze zur Unterstützung der Aussterbeereignisse.
a–d, Daten beziehen sich auf das Timing (a), die Umgebung (b) und das Verhalten (c,d), die von den Standorten präsentiert werden. a, Modellierte Altersbereiche jeder Höhle (n = 22 Höhlen) unter Verwendung des minimalen und maximalen Alters der fossilführenden Einheit (n = 157 Proben). Die Höhlen (x-Achse) im Vergleich zum Alter (y-Achse), mit G. blacki (grüne Kreise) und Nicht-G. Blacki (orangefarbene Kreise) Brekzie. Die Datenpunkte stellen Durchschnittsalter mit Standardabweichung dar. bei 2σ Unsicherheiten. Die Einschübe sind modellierte Brekzien aus Queque (i) und Baxian (ii). G, G. blacki-führende Brekzie; F1, darüber liegender Fließstein; und Non-G, Abwesenheit von G. blacki. Datenpunkte sind Durchschnittsalter mit Standardabweichung. bei 2σ Unsicherheiten. Die schwarzen horizontalen Rechtecke (mit gestrichelten Linien) stellen die Grenze gemäß der Modellierung dar (Ergänzende Informationen, Abschnitt 2 und Ergänzende Abbildung S1a – v). Die modellierte EW ist die vertikale graue Linie. b, Prozentsatz der Pollen von den Standorten in a, die Baumarten (grün), Nichtbaumarten (gelb) und Farne (orange) repräsentieren. Die Kreisdiagramme liefern einen Durchschnitt der Pollenveränderungen vor (links) und nach dem Aussterben (rechts). c, DMTA-Boxplot-Reihe nach Alter von 12 Höhlen (x-Achse) versus molarer Mikroverschleißkomplexität (Asfc, oben, y-Achse) und Anisotropie (epLsar, unten, y-Achse) von G. blacki (rot, n = 16) und P . weidenreichi (blau, n = 22). Die Größenbereiche der Boxplots stellen mittlere Komplexitäts- und Anisotropiewerte pro Standort dar. Die Daten werden als Mittelwerte ± Interquartilbereich und Whiskers bei 95 % KI dargestellt (Ergänzungstabelle S28). d, Spurenelementkartierung von G. blacki und P. weidenreichi. Sr/Ca (i) und Ba/Ca (ii) eines rechten M3 G. blacki-Zahns (CSQSN-44) und Sr/Ca-Karte eines rechten M2 P. weidenreichi-Zahns (CSQ0811-4) (iii), alle aus Queque Höhle. Unten: Sr/Ca (iv) und Ba/Ca (v) aus einem P4-Zahn von G. blacki (ST_02_109) im Vergleich zu Sr/Ca (vi) aus einem linken M3-Zahn von P. weidenreichi (CLMST0911-118), alle aus Shuangtan-Höhle. a.u., beliebige Einheiten.
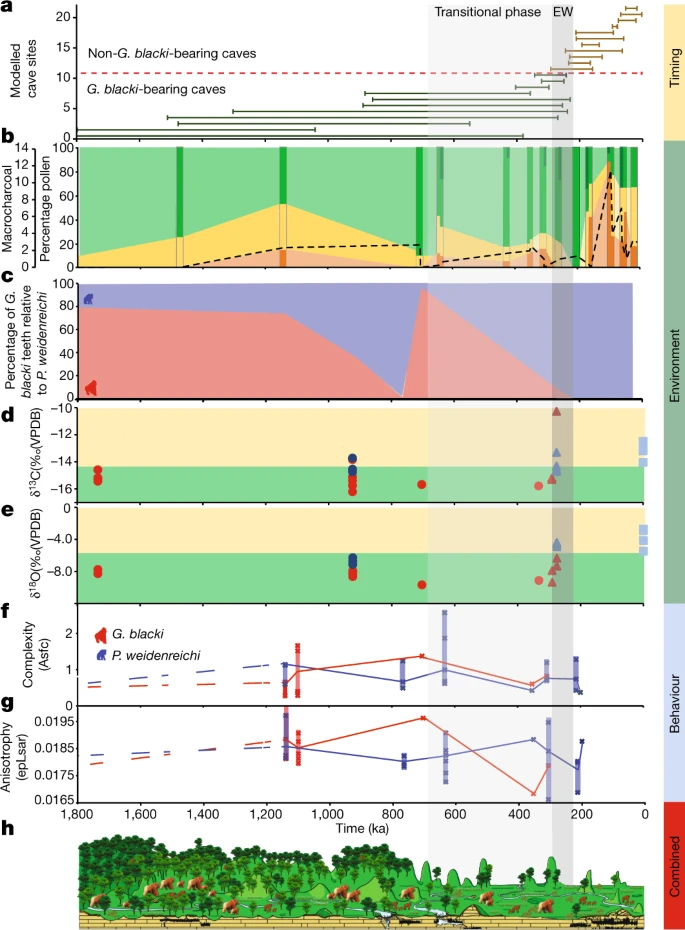
Abb. 3: Eine Zusammenfassung aller Datensätze, chronologisch aufbereitet.
a, Zeitleiste des Aussterbens basierend auf den modellierten Altersspannen für alle 22 Höhlen. Die Zahlen auf der y-Achse beziehen sich auf die Höhlen in Abb. 2a. Beachten Sie die verkürzte Zeitleiste (1.800 ka). Die EW (255 ± 40 ka) ist eine vertikale graue Box (EW) mit einer durchgezogenen, hellgrauen Box (Übergangsphase) für den Beginn einer erhöhten Umweltvariabilität. b, Der Prozentsatz an Pollen, aufgetragen auf einer Zeitachse, gruppiert in baumartige (grün), nicht baumartige (gelb) und Farne (orange). Die dunkleren Streifen stellen Standorte dar, die Pollendaten enthalten, während die helleren Abschnitte dazwischen eine Schätzung der Pollenveränderungen darstellen. Die Mikrokohle (schwarze gestrichelte Linie) korreliert mit der Zunahme der Farne und dem Rückgang der Baumbedeckung. Die dunkelgrünen Baumabschnitte stellen Waldstörungen dar. Taxa mit hohem Umsatz wie Trema, Celtis und Sapindaceae sind während der Übergangsphase und in Ostwesteuropa vorhanden. c, Der Prozentsatz der Zähne von G. blacki (rot) im Verhältnis zu den Zähnen von P. weidenreichi (blau) in repräsentativen Höhlen als grober Indikator für die relative Häufigkeit von G. blacki im Vergleich zu P. weidenreichi an jedem Standort. Die relative Anzahl der G. blacki-Zähne nimmt unmittelbar vor der Übergangsphase ab, was eine Veränderung in der Zusammensetzung der Fauna darstellt, und während der Übergangsphase, was die Ausrottung von G. blacki bedeutet. d,e, Isotopenänderungen für fossile Zähne von P. weidenreichi (blaue Kreise und Dreiecke) und G blacki (rote Kreise und Dreiecke), aufgezeichnet auf einer Zeitachse; moderne P. weidenreichi sind blaue Quadrate. δ13C (‰) (d) und δ18O(‰) (e). f,g, DMTA-Boxplot-Zeitreihen für Mikroverschleißkomplexität (f) und Anisotropie (g) von G. blacki (rot) und P. weidenreichi (blau); Definitionen siehe Abb. 2c. h, Ein Landschafts- und Umweltzeitschnitt, der die Veränderung der Vegetation und der Primatenarten von der Zeit vor der EW über die EW bis zur Zeit nach der EW zeigt.
Dir durchgeführte Pollenanalyse zeigt, daß die Umwelt während der Vor-EW von Baumarten (Pinaceae, Fagaceae und Betulaceae) mit Grünlandflächen dominiert wurde (Abb. 2b und 3b). Allerdings kam es vor der EW während der Übergangsphase zu einer Veränderung der Waldpflanzengemeinschaften und einer Zunahme der Waldstörungstaxa, wobei offenere Wälder dominierten. Nach der EW etwa 200 ka gab es einen starken Rückgang der Baumbedeckung, eine Zunahme der Farne (z. B. Moraceae und Podocarpus), eine starke Zunahme des Graslandes (z. B. Poaceae) und vermehrte Anzeichen von Holzkohle in der Landschaft (Erweitert). Daten Abb. 7 und ergänzende Informationen Abschnitt 10).
Eine detaillierte Faunenanalyse zeigt, daß die Standorte vor der EW durch G. blacki (in relativ großer Zahl) (Abb. 3c), Ailuropoda microta, Procynocephalus, Sinomastodon, Stegodon, Hesperotherium und Hippopotamodon gekennzeichnet waren, die sich zu G. blacki (in relativ großer Zahl) verlagerten kleine Zahlen) (Abb. 3c), Ailuropoda baconi, Stegodon und Elephas vor dem EW und ein Fehlen von G. blacki nach dem EW (Ergänzende Informationen, Abschnitt 3). Die mikrostratigraphischen Analysen von fünf Höhlen zeigen vor-EW-Mikrofazies, die von feinen Körnern, höheren Tonen und Oxiden, Bioturbation und Guano-induzierter Phosphatierung dominiert werden. Im EW nahmen die Korngrößen zu, wobei geringere Oxide, Bioturbation und Knochen-/Zahnveränderungen eine bessere Fossilerhaltung ermöglichten. Während der Zeit nach der EW kehrte dies zu den Merkmalen vor der EW zurück (Erweiterte Daten, Abb. 8c und Abschnitt 11 mit ergänzenden Informationen).
Die stabilen Isotopendaten deuten darauf hin, dass δ13C und δ18O von G. blacki für die Zeit vor dem EW zwischen −16,2 und −13,8 ‰ bzw. −9,7 und −7,0 ‰ liegen. Während der EW steigt dieser leicht auf −15,3 bis −10,3 ‰ bzw. −9,3 bis −6,3 ‰. Im Fall von P. weidenreichi sind die δ13C- und δ18O-Bereiche vor EW ähnlich bei –14,7 bis –13,7‰ und –7,1 bis –6,3‰, erstrecken sich auf –14,7 bis –13,3‰ und ändern sich auf –4,9 und –4,4‰ während der EW-Periode (Abb. 3d, e, Erweiterte Daten Abb. 8b und Abschnitt 12 mit ergänzenden Informationen).
Die Spurenelementanalyse der G. blacki-Zähne vor der EW zeigt mehrere deutliche und synchrone Sr/Ca- und Ba/Ca-Streifen im Zahnschmelz und Dentin, die sich in deutlich weniger sichtbare diffuse Streifen näher an der EW ändern (Abb. 2d, erweitert). Daten Abb. 9 und 10a und ergänzende Informationen Abschnitt 13). Darüber hinaus ist in der Prä-EW eine deutliche Ableitungsstreifenbildung zu erkennen, die während der EW weniger deutlich wird (Extended Data Abb. 10a). Die Microwear-Analyse zeigt keine statistisch signifikanten Ernährungsunterschiede zwischen G. blacki- und P. weidenreichi-tragenden Standorten (Ergänzende Informationen, Abschnitt 14). Es gibt jedoch erhebliche Unterschiede in der Ernährung an vier G. blacki-tragenden Standorten zwischen der Zeit vor der EW und kurz vor der EW. G. blacki neigt dazu, etwas höhere Schwankungen der mittleren Anisotropie- und Komplexitätstrendlinien zu zeigen, wohingegen die von P. weidenreichi stabiler zu sein scheinen, insbesondere für die Anisotropie über die EW hinaus (Abb. 2c und 3f, g, erweiterte Daten Abb. 10b und). Ergänzende Informationen Abschnitt 14).
Zum ersten Mal wurde die größte Sammlung von In-situ-Beweisen von G. blacki über sein gesamtes Verbreitungsgebiet genau datiert, um eine genaue Zeitleiste für die Anwesenheit und Abwesenheit von G. blacki im Fossilienbestand zu liefern. Frühere Datierungen konzentrierten sich hauptsächlich auf die früheren Beweise für G. blacki2,8 und ortsspezifische Chronologien (z. B. Lit. 9). Im Gegensatz dazu wurde durch die Eingrenzung von Höhlen innerhalb des gesamten Altersbereichs sowohl im Chongzuo- als auch im Bubing-Becken ein genaueres regionales Fenster des Aussterbens bei 295–215 ka ermittelt.
Die Pollen- und Faunendaten deuten darauf hin, daß die frühen Mosaiklandschaften vor der EW in der Übergangsphase durch eine erhöhte Umweltvariabilität unterbrochen wurden (Abb. 3b), was durch die Veränderung der Waldgemeinschaften und -strukturen nahegelegt wird, und nach der EW, wie durch einen Rückgang der Baumbestände nahegelegt Bedeckung und eine Zunahme von Farnen und Grasland im Zusammenhang mit Bränden. Diese Variabilität begann schrittweise zwischen 1.100 und 350 ka und nahm ab etwa 200 ka dramatisch zu (Abb. 3b). Es wurde diese Variabilität als Verschiebungen hin zu erhöhter Saisonalität und trockeneren Umgebungen interpretiert, die zu einer Verlagerung hin zu saisonalen subtropischen/tropischen feuchten Tieflandwäldern und einer Zunahme von Sträuchern und offenen Graslandumgebungen vor und während der EW führten (Ergänzende Informationen, Abschnitt 10). Diese Umweltvariabilität zeigt sich auch in der Sedimentaufzeichnung, da die stabilen Niedrigenergieumgebungen der Vor-EW durch instabile Hochenergieumgebungen der EW ersetzt wurden, wobei die Wasserverfügbarkeit auf die Regenzeit beschränkt war (Erweiterte Daten, Abb. 8c und Abschnitt „Ergänzende Informationen“) 11).
Der Rückgang der Waldfläche in diesem Zeitraum ist in China21, Südostasien22 und Australasien23 dokumentiert. Die Pollenstudie zeigt jedoch, daß der Schlüssel zum Aussterben von G. blacki nicht in der Verschlechterung der Baumbedeckung liegt, sondern vielmehr im Einfluss von Umweltschwankungen auf die Veränderung der Zusammensetzung von Waldgemeinschaften, insbesondere in der Zunahme von Störtaxa. Die gewonnenen stabilen Isotopen- und Spurenelementdaten liefern neue Einblicke in das Ausmaß dieser Variabilität und die Auswirkungen auf G. blacki (Ergänzende Informationen, Abschnitte 12 und 13). Vor EW lebten G. blacki und P. weidenreichi beide in bewaldeten Umgebungen mit geschlossenen Baumkronen (Abb. 3b und erweiterte Daten, Abb. 8b) mit stärkerer biogener Streifenbildung (Abb. 2d(i)–(iii)), was wahrscheinlich auf a zurückzuführen ist größere Vielfalt an Nahrungsquellen, einschließlich saisonaler Früchte und Blumen und periodischer Wasserkonsum, wie durch die deutliche Bleistreifenmarkierung angezeigt (Erweiterte Daten Abb. 10a,b). Die wahrscheinlichsten Nahrungsquellen wären das ganze Jahr über in größerer Verfügbarkeit gewesen und hätten nur diskreten Stress in der Bevölkerung verursacht (Abb. 2d(i)–(iii)). Mit Ausnahme eines Individuums scheint G. blacki während der gesamten EW-Periode eine spezialisiertere geschlossene Baumkronennische unterhalten zu haben, die möglicherweise auf eine Mischung von Waldpflanzen angewiesen ist (Erweiterte Daten, Abb. 8b). Diese Spezialisierung während einer Umweltveränderung könnte zu einem diffuseren biogenen Signal im Zahngewebe der einzelnen Personen geführt haben (Abb. 2d(iv)–(v)), was auf eine stark verringerte Ernährungsvielfalt und einen geringeren regelmäßigen Wasserverbrauch schließen lässt (Erweiterte Daten, Abb. 10c). ,d) und erhöhter chronischer Stress in der Bevölkerung (Abb. 2d(iv)–(v)). Dies ist der erste Einblick in das Verhalten von G. blacki als einer Art am Rande des Aussterbens, was im krassen Gegensatz zu P. weidenreichi (Abb. 2d(vi)) steht, das zu diesem Zeitpunkt viel weniger Stress zeigt. Über den EW hinaus scheint sich P. weidenreichi verlagert zu haben, um die offeneren, saisonalen Lebensräume zu nutzen (Extended Data Abb. 8b) und möglicherweise weiterhin die saisonale Befruchtung von Früchten zu nutzen, wie es das moderne Pongo heute auf Borneo tut24.
Die Veränderungen der Mikroverschleißwerte bei G. blacki- und P. weidenreichi-Zähnen können auch mit Perioden der Fruchtknappheit zusammenhängen. G. blacki neigt dazu, spezifischere Ernährungspräferenzen zu zeigen (sowohl bei Früchten als auch bei faserhaltigen Nahrungsmitteln), was auf eine stärkere Abhängigkeit von faserhaltigen Ersatznahrungsmitteln hinweist (Abb. 2c), beispielsweise in der Ostwestküste, als das Klima saisonaler wurde und weniger Früchte verfügbar waren. Dies könnte G. blacki dazu gezwungen haben, seine Ernährung von ernährungsphysiologisch bevorzugten Komponenten in geringerem Angebot auf weniger nährstoffreiche Ersatznahrungsmittel in reichlichem Angebot umzustellen. Der erhöhte Verzehr von faserhaltigen Lebensmitteln bei P. weidenreichi im EW könnte auf eine bessere Umstellung auf Ersatznahrungsmittel und eine insgesamt flexiblere und ausgewogenere Ernährung hinweisen (Abb. 2c und erweiterte Daten, Abb. 10). Diese erste DMTA-Analyse des gesamten G. blacki-Materialspektrums bietet einen einzigartigen Einblick in seine Unfähigkeit, sich anzupassen, und seine möglicherweise schlechte Auswahl an Ersatznahrungsmitteln.
Diese Studie präsentiert einen genauen Zeitplan für das Vorkommen und Aussterben von G. blacki. In der Zeit vor der EW blühte G. blacki zusammen mit anderen Primaten als erfolgreicher Spezialist auf (Abb. 3c) und genoss eine große Nahrungsvielfalt in einem reichen immergrünen Laubwald (Abb. 2d(i)–(ii)) und reichlich Wasserquellen (Erweiterte Daten Abb. 10a–d) unter stabilen Umweltbedingungen (Abb. 2b). Etwa vor 700–600 ka kam es in der Übergangsphase zu einer Verschiebung hin zu einer zunehmenden Saisonalität, die zu einer Veränderung der Waldgemeinschaften (Abb. 3b), einer geringeren Vielfalt an Nahrungsquellen (Abb. 2d(iii)–(iv)) und instabilen hohen Temperaturen führte. Energieumgebungen (Erweiterte Daten, Abb. 8c), Veränderungen in der Zusammensetzung der Fauna und weit verbreitete Faunenumsätze (Abb. 3c und ergänzende Informationen, Abschnitt 2), eine Verlagerung hin zu saisonalen Lebensräumen durch P. weidenreichi (Erweiterte Daten, Abb. 8b) und a Veränderung der Ernährungsvielfalt und des Verhaltens von G. blacki (Abb. 2d und 3f, g).
Obwohl P. weidenreichi vor der EW eine ähnliche Umgebung hatte, gibt es zwischen 600 und 300 Jahren Hinweise darauf, daß G. blacki nicht in der Lage war, sich an diese Übergangszeit anzupassen, was einen größeren Einfluss auf seine Widerstandsfähigkeit gegenüber der sich verändernden Ökologie hatte. Die Abhängigkeit von G. blacki von Früchten und nährstoffärmeren Ersatznahrungsmitteln (Abb. 2c) führte zu einer risikoreicheren Futtersuchstrategie und machte ihn in Kombination mit seiner viel größeren, weniger beweglichen Körpergröße anfälliger für Veränderungen in der Waldstruktur25 (Abb . 2c). Darüber hinaus war G. blacki ausschließlich terrestrisch, möglicherweise mit einem kleinen geografischen Verbreitungsgebiet20, reiste jedoch regelmäßig das Tal hinunter, um Wasser zu verbrauchen (Erweiterte Daten Abb. 10a–d), wohingegen P. weidenreichi eher baumbewohnend, mobil und halbsolitär war und Wasser im Blatt sammelte Überdachung. Darüber hinaus deuten die einzigartigen dentognathen Merkmale13,14 und die riesige Körpergröße4,5 von G. blacki auf einen höheren Bedarf an Nahrungsaufnahme und ein langsameres und verzögerteres Wachstumsmuster hin, was auf eine geringere Reproduktionsrate schließen lässt26. Obwohl die Zahngröße von G. blacki im Laufe des Pleistozäns zunahm, was auch eine Zunahme der Körpergröße bedeutet, nahm die Größe von P. weidenreichi ab27. Dies macht ihn zu einem agileren Adapter. P. weidenreichi zeigte auch eine Flexibilität gegenüber offenen Lebensräumen (Extended Data Abb. 8b), die sich möglicherweise in kleineren Gruppen bewegten, und war in der Lage, sein Verhalten als Reaktion auf Umweltschwankungen anzupassen, was zu einer weniger gestressten Population führte (Abb. 3d).
Vor etwa 300 Jahren gibt es Hinweise auf eine kämpfende G. blacki-Population, da die Anzahl der G. blacki-Höhlen und -Zähne abnahm (Abb. 3c), was auf eine schrumpfende Population hindeutet. Die starke Veränderung der Zahnbänder von G. blacki weist auf chronischen Stress in der Bevölkerung hin (Abb. 2d(iv)–(v)) und weist auf Veränderungen des bevorzugten Ernährungsverhaltens (Abb. 2c und Extended Data Abb. 10f, g) hin dass G. blacki Schwierigkeiten hatte, auf die Umweltveränderungen in einem potenziell schrumpfenden Gebiet zu reagieren20. Es scheint, daß seine Waldrefugien ihre Struktur verändert haben und zu offen und gestört geworden sind, als dass sich diese Art ernähren könnte. Im Vergleich zu anderen bekannten Aussterbeereignissen in Nordamerika und Australien, die vom Homo sapiens28,29,30 beeinflusst wurden, gibt es keine Hinweise darauf, dass archaische Homininen bei diesem früheren Aussterben der Megafauna in Südchina eine Rolle gespielt haben.
Die Angabe einer definierten Ursache für das Aussterben ist für viele ausgestorbene Arten eine Leistung, die selten erreicht wurde, da sie einen gattungs- und artspezifischen Ansatz erfordert28. Obwohl es sehr schwierig sein kann, die genauen Ursachen für die Ausrottung und das Aussterben der Megafauna zu bestimmen29,30, liefert unsere Multiproxy-Aufzeichnung des Zeitpunkts, der Umwelt und des Verhaltens von G. blacki fundierte regionale Einblicke in den ökologischen Kontext dieser Art. G. blacki war der ultimative Spezialist und als sich die Baumumgebung veränderte, besiegelte sein Kampf um die Anpassung sein Schicksal. Im Vergleich dazu verbreitete und diversifizierte sich der generalistische Homo in dieser Zeit über ganz Südostasien und schien die neuen Mosaikumgebungen, die für G. blacki ein solches Problem darstellten, flexibel ausgenutzt zu haben. Insgesamt liefert unser Datensatz einen wichtigen Kontext für die sich verändernden Schicksale verschiedener Primatenarten in Südostasien und wirft neues Licht auf den Untergang des größten Primaten, der jemals auf dem Planeten gelebt hat.
Samuel Boury, Scott Weady und Leif Ristroph Physik. Rev. Fluids 8, 110503 – Veröffentlicht am 16. November 2023
Einige geologische Beweise deuten darauf hin, daß die Große Sphinx eine natürliche Landform war, bevor ihre Oberfläche von den alten Ägyptern verändert wurde [1,2]. Ist diese umstrittene Theorie überhaupt plausibel? Etwas Unterstützung kommt von einer Klasse von Landformen namens Yardangs, die sitzenden Löwen ähneln, aber wie und warum sie solche Formen annehmen, ist rätselhaft [3,4]. Strömungsmechanische Untersuchungen können Erkenntnisse liefern, indem sie zeigen, welche Arten von Formationen durch Strömungserosion geformt werden können, und Visualisierungsstudien können die Mechanismen aufdecken, die die beobachteten Formen mit den Strömungen verbinden.
Die Wissenschaftler untersuchten dieses Problem, indem sie Laborexperimente zur Erosion von Körpern aus Ton durchführten, die von schnell fließendem Wasser umspült werden. Basierend auf Berichten über die ungleichmäßige Zusammensetzung des Gesteins, aus dem die Sphinx besteht [5], haben sie die Wirkung harter, nicht abbaubarer Einschlüsse in Hügeln aus weicherem Ton getestet. Als Idealisierung eines vorherrschenden Windmusters setzten sie die Körper der unidirektionalen Strömung eines Wassertunnels aus. Unter geeigneten Bedingungen stellen sie fest, daß sphinxartige „Skulpturen“ durch die Strömung geformt werden, wie in den Abbildungen 1 und 2 gezeigt.
Abbildung 1
Eine Laborsphinx in der Strömung eines Wassertunnels.Das Objekt ist mit Ton und Fluoreszeinfarbstoff beschichtet, und das Foto fängt das „Streifenvolumen“ oder den 3D-Flussbereich ein, der irgendwann in die Grenzschicht eingedrungen ist und die Oberfläche erodiert hat.
Abbildung 2 Visualisierung von Strömungsstreifenlinien in vertikaler (links) und horizontaler (rechts) Ebene. Nicht dargestellt ist eine Reihe von Injektionsschläuchen, die Filamente aus Fluorescein-Farbstoff in den stromaufwärts gelegenen Strom abgeben.
Unter der Annahme, daß die ursprüngliche Form keine Merkmale aufweisen sollte, bauten sie einen Hügel aus Bentonit-Ton zu einem halben Ellipsoid auf, dessen Längsachse mit der Strömung aus einem Wassertunnel ausgerichtet ist (Engineering Laboratory Design). Der Ton ist ein Pulver, dem Wasser im Verhältnis 2:1 zugesetzt wird, um eine steife Paste zu bilden, die Schicht für Schicht auf eine Plattform aufgetragen wird, die als „Grundgestein“ dient. Die Materialinhomogenität hat die einfache Form eines kurzen, erosionsbeständigen Kunststoffzylinders, der zunächst vollständig im Ton eingeschlossen ist. Das Foto in Abb. 1 zeigt ein späteres Stadium, das einem ruhenden Löwen ähnelt. Der inzwischen weitgehend ausgegrabene zylindrische Einschluss ist zum „Kopf“ geworden, der dem Wind zugewandt ist. Es ist unterschnitten, um einen „Hals“ zu bilden, der mit dem Körper verbunden ist, während „Pfoten“ an der Basis verbleiben. Was verursacht diese Anatomie?
Die Laborumgebung komprimiert Zeit und Raum im Vergleich zu dem, was in der Natur geschieht. Die laborgemachten Strukturen entwickeln sich über Stunden und ihre Größe ermöglicht es den Forschern, die 3D-Morphologie in regelmäßigen Abständen mit einem optischen Scanner (Shinning 3D EinScan-SE) zu erfassen. Aus der Strömung entfernt, wird der Körper mit Lichtmustern beleuchtet und aus vielen Winkeln stereoskopisch abgebildet. Diese Daten fließen in eine digitale Rekonstruktion mit einer Auflösung von etwa 0,1 mm ein. Da die Tonskulptur vergänglich ist, eignet sie sich schlecht für die Strömungsvisualisierung. Stattdessen wandeln die Forscher den Oberflächenscan im Sphinx-ähnlichen Stadium in eine Datei mit fester Geometrie um, die mithilfe eines Stereolithographiedruckers (Formlabs Form 3L) in 3D aus Kunststoffharz gedruckt wird.
Das Foto in Abb. 1 entsteht, indem eine dünne Schicht aus mit Fluoreszeinfarbstoff vermischtem Ton über das gedruckte Formular aufgetragen wird [6], das dann zur Aufnahme mit einer Digitalkamera (Nikon D610) in den Strömungstunnel zurückgeführt wird. Die neongrünen Farbtöne des Originalbildes werden in Sand- und Ockertöne umgewandelt. Ein hoher Kontrast wird durch mattschwarze Hintergrundplatten und die Beleuchtung durch hellweiße Lampen erzielt, die darauf abzielen, die Farbe zu verstärken, ohne Schatten zu werfen. Die Verwendung eines vergrößerten Körpers erwies sich als nützlich, um die gleiche Reynolds-Zahl Re bei geringerer Strömungsgeschwindigkeit zu erreichen, was zu einer geringeren Dispersion des Farbstoffs führt. Eine niedrigere Geschwindigkeit ist auch hilfreich, um die Arbeitszeit zum Sammeln von Fotos zu verlängern, die im Umlauftunnel durch die Rückführung der verschmutzten Flüssigkeit in den Testabschnitt begrenzt ist.
Abbildung 1 kann als „Streifenvolumen“ oder als 3D-Bereich interpretiert werden, der von der gesamten Flüssigkeit ausgeschwemmt wird, die innerhalb der Grenzschicht geflossen ist und die Oberfläche erodiert hat. Offensichtlich ist ein Großteil des Körpers von getrennten Strömungen und dem turbulenten Wirbelstrom umgeben. Vom Kopf austretende Wirbel bilden die wellige und wogende „Mähne“ des Löwen, und diese Strömungen scheinen für die verstärkte Erosion unmittelbar stromabwärts des Kopfes verantwortlich zu sein, die zur Entstehung des gewölbten Rückens führt.
Die Luv-Merkmale des Löwen lassen sich besser durch Streifenlinienbildgebung mit Farbstofffäden erklären, die vor der Struktur freigesetzt werden, wie in den Seiten- und Draufsichtfotos von Abb. 2 gezeigt. Hier wurden Anordnungen von Injektionsschläuchen verwendt, durch die hindurch Fluorescein-Farbstoff wird zugeführt und die Bilder werden erneut umgefärbt. Um gerade statt gewundene Filamente zu erhalten, sind schlanke Rohre erforderlich, deren Enden sorgfältig entgratet und abgeschrägt sind und die sanft gebogen werden, um sich sanft an die Strömung anzupassen. Ein größeres Modell und eine langsamere Strömungsgeschwindigkeit tragen wiederum dazu bei, die Dispersion des Farbstoffs zu minimieren. Es ist unnötig, aber hilfreich, die Dichte des Farbstoffs durch Zugabe von Alkohol und die Fließgeschwindigkeit anzupassen, was wir mit einer Mariotte-Flasche erreichen, die auch eine konstante Zufuhrrate aufrechterhält.
Es wird beobachtet, daß die Teile der Streifenlinien direkt stromaufwärts und an den Seiten der Kopfregion stabil sind. Sie können daher als Stromlinien interpretiert werden, deren Abstände die lokale Strömungsgeschwindigkeit anzeigen, wobei enger beieinander liegende Linien aufgrund der Massenerhaltung des inkompressiblen Fluids mit einer schnelleren Strömung einhergehen. Das Seitenansichtsbild zeigt schnellere Strömungen im Bereich des Halses des Löwen, da der Strom vom Kopf oben und den Pfoten unten eingeleitet wird. Das Draufsichtbild zeigt auch schnellere Strömungen in diesem Bereich, was in der horizontalen Ebene auf die Aufteilung und Umlenkung des einströmenden Stroms um den Hals zurückzuführen ist. Diese verstärkenden Effekte könnten die lokal hohe Scherbeanspruchung und die hohe Erosionsrate direkt unter dem Kopf erklären und damit erklären, warum starkes Schnitzen den Hals eingräbt und die Pfoten freilegt.
Das Laborsystem ist bestenfalls von Landformen inspiriert und hat einen qualitativen Bezug zu ihnen. Es wurden Objekte mit einer typischen Größe von 10 cm in Wasserströmungen mit einer typischen Geschwindigkeit von 10 cm/s untersucht und ergeben Re=O(104), die um Größenordnungen niedriger sind als die, die für natürliche Yardangs auftreten. Ton in fließendem Wasser gehorcht einem einfachen Gesetz, bei dem die Erosionsrate mit der lokalen Fluidscherspannung auf der festen Oberfläche variiert [7,8]. Im Gegensatz dazu umfassen die relevanten äolischen oder windgetriebenen Prozesse den Abrieb durch vom Wind getragene Körner und die Abreibung oder den Transport gelöster Körner durch turbulente Strömungen [9,10]. Dennoch sind möglicherweise einige allgemeine Aspekte des Problems der Formflussentwicklung robust gegenüber Systemdetails.
Die Forschungsergebnisse legen nahe, daß sich unter recht alltäglichen Bedingungen sphinxartige Strukturen bilden können. Diese Erkenntnisse lösen kaum die Geheimnisse hinter Yardangs und der Großen Sphinx, aber vielleicht regen sie uns zu der Frage an, auf welche beeindruckenden Landformen die alten Völker in den Wüsten Ägyptens gestoßen sein könnten und warum sie sich ein phantastisches Geschöpf vorgestellt haben könnten.
Wir danken R. Mehta und K. Long vom NASA Ames Research Center für Diskussionen über Streak-Line-Bildgebung und danken der National Science Foundation für ihre Unterstützung durch den Zuschuss Nr. DMS-2206573.
Referenzen 1 F. El-Baz, Desert builders knew a good thing when they saw it, Smithsonian 12, 116 (1981). 2 G. Gerster and F. El-Baz, Egypt’s desert of promise, National Geographic 161, 190 (1982). 3 A. S. Goudie, Mega-yardangs: A global analysis, Geography Compass 1, 65 (2007). 4 A. J. Parsons and A. D. Abrajams, Geomorphology of Desert Environments, 2nd ed. (Springer, Dordrecht, 2009). 5 K. L. Gauri, Geologic study of the sphinx, Newsletter of the American Research Centre In Egypt 127, 24 (1984). 6 K. Amin, J. M. Huang, K. J. Hu, J. Zhang, and L. Ristroph, The role of shape-dependent flight stability in the origin of oriented meteorites, Proc. Natl. Acad. Sci. USA 116, 16180 (2019). 7 M. N. J. Moore, L. Ristroph, S. Childress, J. Zhang, and M. J. Shelley, Self-similar evolution of a body eroding in a fluid flow, Phys. Fluids 25, 116602 (2013). 8 L. Ristroph, M. N. J. Moore, S. Childress, M. J. Shelley, and J. Zhang, Sculpting of an erodible body by flowing water, Proc. Natl. Acad. Sci. USA 109, 19606 (2012). 9 R. S. Anderson, Erosion profiles due to particles entrained by wind: Application of an eolian sediment-transport model, Geological Society of America Bulletin 97, 1270 (1986). 10 M. S. Yalin, Mechanics of Sediment Transport (Pergamon, Oxford, 1976).
Die Geschwindigkeit, mit der sich das Universum ausdehnt, bekannt als Hubble-Konstante, ist einer der grundlegenden Parameter für das Verständnis der Entwicklung und des endgültigen Schicksals des Kosmos. Es besteht jedoch ein anhaltender Unterschied, der als „Hubble-Spannung“ bezeichnet wird, zwischen dem Wert der Konstante, der mit einer Vielzahl unabhängiger Entfernungsindikatoren gemessen wird, und dem Wert, der aus dem Nachglühen des Urknalls vorhergesagt wird.
Kombinierte Beobachtungen der NIRCam (Near-Infrared Camera) der NASA und der WFC3 (Wide Field Camera 3) von Hubble zeigen die Spiralgalaxie NGC 5584, die 72 Millionen Lichtjahre von der Erde entfernt liegt. Zu den leuchtenden Sternen von NGC 5584 gehören pulsierende Sterne, sogenannte Cepheid-Variablen, und Supernovae vom Typ Ia, eine besondere Klasse explodierender Sterne. Astronomen nutzen Cepheid-Variablen und Typ-Ia-Supernovae als zuverlässige Entfernungsmarker, um die Expansionsrate des Universums zu messen. Credit: NASA, ESA, CSA, and A. Riess (STScI).
Das James-Webb-Weltraumteleskop der NASA bietet neue Möglichkeiten, einige der stärksten Beobachtungsbeweise für diese Spannung zu untersuchen und zu verfeinern. Nobelpreisträger Adam Riess von der Johns Hopkins University und dem Space Telescope Science Institute stellt die jüngste Arbeit von ihm und seinen Kollegen vor, bei der er Webb-Beobachtungen nutzte, um die Präzision lokaler Messungen der Hubble-Konstante zu verbessern.
„Hatten Sie jemals Schwierigkeiten, ein Schild zu erkennen, das sich am Rande Ihres Sichtfelds befand? Was sagt es? Was bedeutet das? Selbst mit den leistungsstärksten Teleskopen erscheinen die „Zeichen“, die Astronomen lesen wollen, so klein, dass auch wir Schwierigkeiten haben.
„Das Zeichen, das Kosmologen lesen wollen, ist ein kosmisches Geschwindigkeitsbegrenzungszeichen, das uns sagt, wie schnell sich das Universum ausdehnt – eine Zahl, die Hubble-Konstante genannt wird.“ Unser Zeichen ist in die Sterne entfernter Galaxien eingeschrieben. Die Helligkeiten bestimmter Sterne in diesen Galaxien verraten uns, wie weit sie entfernt sind und wie lange dieses Licht somit gereist ist, um uns zu erreichen, und die Rotverschiebungen der Galaxien verraten uns, wie stark sich das Universum in dieser Zeit ausgeweitet hat, und verraten uns damit die Expansionsrate. „Eine besondere Klasse von Sternen, die Cepheid-Variablen, liefert uns seit über einem Jahrhundert die genauesten Entfernungsmessungen, weil diese Sterne außerordentlich hell sind: Es handelt sich um Überriesensterne mit der hunderttausendfachen Leuchtkraft der Sonne.“ Darüber hinaus pulsieren sie über einen Zeitraum von Wochen (das heißt, sie vergrößern und verkleinern sich), was ihre relative Leuchtkraft anzeigt. Je länger die Periode, desto heller sind sie. Sie sind das Goldstandardwerkzeug zur Messung der Entfernungen von Galaxien, die hundert Millionen oder mehr Lichtjahre entfernt sind, ein entscheidender Schritt zur Bestimmung der Hubble-Konstante. Leider sind Sterne in Galaxien von unserem entfernten Standpunkt aus auf engstem Raum zusammengedrängt und daher fehlt uns oft die Auflösung, sie von ihren Nachbarn in der Sichtlinie zu trennen.
„Eine wichtige Begründung für den Bau des Hubble-Weltraumteleskops war die Lösung dieses Problems. Vor dem Hubble-Start im Jahr 1990 und den anschließenden Cepheid-Messungen war die Expansionsrate des Universums so ungewiss, dass die Astronomen nicht sicher waren, ob sich das Universum seit 10 oder 20 Milliarden Jahren ausdehnt. Das liegt daran, dass eine schnellere Expansionsrate zu einem jüngeren Alter des Universums führt und eine langsamere Expansionsrate zu einem höheren Alter des Universums. Hubble verfügt über eine bessere Auflösung im sichtbaren Wellenlängenbereich als jedes bodengestützte Teleskop, da es sich über den Unschärfeeffekten der Erdatmosphäre befindet. Dadurch kann es einzelne Cepheid-Variablen in Galaxien identifizieren, die mehr als hundert Millionen Lichtjahre entfernt sind, und das Zeitintervall messen, in dem sie ihre Helligkeit ändern.
„Allerdings müssen wir die Cepheiden auch im nahen Infrarotbereich des Spektrums beobachten, um das Licht zu sehen, das den dazwischenliegenden Staub unbeschadet durchdringt. (Staub absorbiert und streut blaues optisches Licht, lässt entfernte Objekte schwach erscheinen und täuscht uns vor, sie seien weiter entfernt als sie sind.) Leider ist Hubbles Rotlichtsicht nicht so scharf wie seine Blaulichtsicht, sodass das Licht der Cepheid-Sterne, das wir dort sehen, mit anderen Sternen in seinem Sichtfeld vermischt ist. Wir können die durchschnittliche Mischungsmenge statistisch auf die gleiche Weise erklären, wie ein Arzt Ihr Gewicht ermittelt, indem er das durchschnittliche Gewicht der Kleidung vom Messwert auf der Waage abzieht. Dies führt jedoch zu einer Verzerrung der Messungen. Die Kleidung einiger Menschen ist schwerer als die anderer.
„Die scharfe Infrarotsicht ist jedoch eine der Superkräfte des James-Webb-Weltraumteleskops. Mit seinem großen Spiegel und seiner empfindlichen Optik kann er das Licht der Cepheiden problemlos und mit geringer Überblendung von benachbarten Sternen trennen. Im ersten Jahr der Webb-Operationen mit unserem General Observers-Programm 1685 sammelten wir Beobachtungen von Cepheiden, die Hubble auf zwei Stufen entlang der sogenannten kosmischen Distanzleiter gefunden hatte. Der erste Schritt besteht darin, Cepheiden in einer Galaxie mit bekannter geometrischer Entfernung zu beobachten, die es uns ermöglicht, die wahre Leuchtkraft der Cepheiden zu kalibrieren. Für unser Programm ist diese Galaxie NGC 4258. Der zweite Schritt besteht darin, Cepheiden in den Wirtsgalaxien der jüngsten Supernovae vom Typ Ia zu beobachten. Die Kombination der ersten beiden Schritte überträgt Kenntnisse über die Entfernung zu den Supernovae, um ihre wahre Leuchtkraft zu kalibrieren. Schritt drei besteht darin, die weit entfernten Supernovae zu beobachten, bei denen die Expansion des Universums offensichtlich ist und durch Vergleich der Entfernungen gemessen werden kann, die aus ihrer Helligkeit und den Rotverschiebungen der Supernova-Wirtsgalaxien abgeleitet werden. Diese Abfolge von Schritten wird als Distanzleiter bezeichnet.
„Wir haben kürzlich unsere ersten Webb-Messungen aus den Schritten eins und zwei erhalten, die es uns ermöglichen, die Entfernungsleiter zu vervollständigen und mit den vorherigen Messungen mit Hubble zu vergleichen (siehe Abbildung). Webbs Messungen haben das Rauschen bei den Cepheid-Messungen aufgrund der Auflösung des Observatoriums drastisch reduziert.“ Wellenlängen im nahen Infrarotbereich. Von einer solchen Verbesserung träumen Astronomen! Auf den ersten beiden Stufen haben wir mehr als 320 Cepheiden beobachtet. Wir haben bestätigt, dass die früheren Messungen des Hubble-Weltraumteleskops genau waren, wenn auch mehr Rauschen. Wir haben mit Webb auch vier weitere Supernova-Kandidaten beobachtet und sehen ein ähnliches Ergebnis für die gesamte Stichprobe.